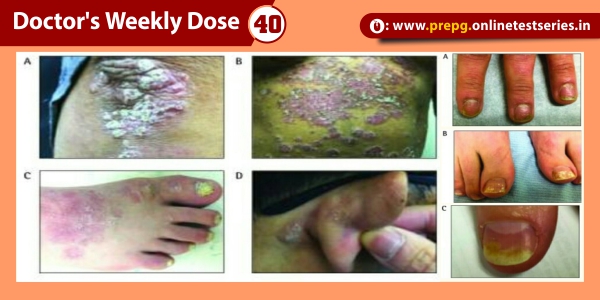
Alpha-1 antitrypsin deficiency, also known as alpha 1-proteinase inhibitor deficiency or, more simply, Alpha-1, is a genetic condition that increases the risk of developing a variety of diseases including pulmonary emphysema and cirrhosis of the liver. It is caused by mutations in the gene coding for the 52 kDa glycoprotein a1-antitrypsin (AAT), the body’s major serine proteinase inhibitor or serpin. This gene is located in the long arm of chromosome 14 of the human genome.
ROLE OF AAT
There does not appear to be a unifying mechanism for the panoply of illnesses associated with AAT deficiency. While AAT is the archetype of the serpin family and is an extremely effective inhibitor of such tissue destroying proteolytic enzymes as neutrophil elastase, it has also been shown to have both anti-and pro-inflammatory properties. The pulmonary emphysema associated with AAT deficiency is due to the unbridled proteolytic activity of neutrophil elastase on lung connective tissue leading to alveolar destruction.
While the proteinase-antiproteinase balance may also play a role in the liver disease of AAT deficiency, most believe that retention of misfolded and polymerised AAT within the endoplasmic reticulum of hepatocytes of affected individuals and the response of hepatocytes to this retained protein are more likely culprits in this form of AAT deficiency-associated disease.
EMERGING NEW TREATMENTS
Alternative routes of administration of current treatment
Most studies are focusing on drug delivery by inhalation. There was initial reluctance to pursue development of inhaled AAT since it was difficult to document that the inhaled protein was able to be delivered to the pulmonary interstitium where proteolytic activity was thought to be doing its damage in pulmonary emphysema. In recent years, however, it has been shown that the airways of individuals with AAT deficiency are under a constant inflammatory barrage and that administration of exogenous inhaled AAT can reconstitute the lower respiratory tract antiproteinase screen and potentially reduce inflammation. Interest in this route of administration has therefore been renewed.
Alternative sources of augmentation therapy
Transgenic production of human AAT protein has been accomplished in both sheep (PPL Therapeutics, Scotland, UK and Bayer Biologicals, West Haven, Conn, USA) and goats (Genzyme, Boston, Mass, USA). Human AAT has also been produced in yeast using recombinant technology (Baxter Healthcare, IL, USA and Arriva Pharmaceuticals, Alameda, CA, USA). Because of anomalies in the glycosylation of the AAT protein in these various species, these proteins are cleared rapidly from the human circulation making intravenous administration impractical. The PPL product and the yeast-derived AAT have been evaluated in human safety studies using an inhaled route of administration.
Treatments aimed at pulmonary emphysema
A number of agents for the treatment of pulmonary emphysema due to smoking are under development.
The first are the retinoids. Studies using elastase-induced emphysema in rats suggested that administration of all-trans retinoic acid (ATRA) was associated with the reversal of the emphysematous changes.
When animals are administered hyaluronic acid they are protected from exogenous elastase-induced emphysema. These findings have led to trials of inhaled hyaluronic acid in individuals with AAT deficiency in the hope of preventing the progression of lung disease.
Finally, our understanding that oxidative inactivation of AAT may lead to loss of antiproteinase activity has led to consideration of drugs with antioxidant potential in the treatment of individuals with AAT deficiency-related emphysema. Physicians have been suggesting the use of supplements containing vitamins A, C, and/or E based on this rationale, and more potent antioxidants are also being considered for evaluation.
Treatments aimed at the liver
Since most deficient genotypes lead to the production of an AAT protein that has fairly potent antineutrophil elastase capacity, one therapeutic concept has been aimed at trying to cause the liver to release its trapped AAT, thus relieving the congestion of the hepatocyte and reconstituting the circulating anti elastase screen. The most promising candidates for this approach are the synthetic chaperones and molecular interventions that try to prevent intracellular polymerisation of the abnormal AAT.
Synthetic chaperones have been used in intracellular protein transport diseases such as cystic fibrosis. The most studied candidate is 4-phenyl-butyric acid (4-PBA) and this has been studied in AAT deficiency at several centres.
Genetic approaches
As a condition caused by well characterised single gene mutations, the possibility of genetic approaches to mitigate or cure AAT deficiency have been entertained. Studies have been reported in which the normal human AAT gene has been inserted into muscle or liver cells. Novel gene repair technologies have also been studied. Additionally, consideration has been given to the development of agents that can turn off or disrupt production of the abnormal gene product.
Animal studies at the University of Florida have succeeded in introducing the normal human AAT gene into striated muscle cells using an adeno-associated virus vector.
More speculative still is the concept of gene repair. This technology was initially designed around chimeraplasty, which is the use of chimeric RNA/DNA oligonucleotides to ‘‘patch’’ a single gene mutation. RNA complementary to the area surrounding the point mutation was synthesised with a contiguous DNA oligonucleotide made of the corrected sequence.